How Do Forests Influence and Stabilize Climate?

HOW DO FORESTS INFLUENCE
AND
STABILIZE CLIMATE?
Paul Kreger, California Resources Agency, 1980
Abstract
Forests stabilize micro and macro climates by:
• Photosynthesizing sunlight through green leaves
• Absorbing carbon dioxide from the air
• Sequestering carbon in standing biomass – roots, stems, branches, and leaves – growing tall canopies for more efficient photosynthesis
• Evaporating water through leaves, dissipating heat to improve photosynthetic efficiency and cool forest areas
• Recycling water vapor across valleys and mountains increasing precipitation in marginal areas
• Minimizing soil erosion with layers of leaf litter and growing interlocking root systems that help stabilize seasonally unstable hills and mountains
• Adding nutrient-rich fallen leaves to soil which becomes humus enabling forest soils to store more water and carbon than most other soils
• Increasing the capacity of a site to support life, building on itself, stabilizing the site and the biota. Mineral nutrients are conserved and recirculated, offering opportunities for more life.
• Building climax communities that are stable, self-regulating, self-healing, and resilient
• Assisting in the essential global recycling of water, oxygen, carbon, and nitrogen — without expending fossil fuels.
• Influencing the wind, temperature, humidity, precipitation, soil, and water and energy flow in ways often discovered only after the forests are cut, and these functions are reduced or lost.
• Maintaining high efficiency, low entropy processes: matter and energy are moved around as little as possible to minimize entropy (waste and disorder), information is moved around as much as possible to maximize synergy (efficiency and cooperation), to minimize entropy (waste and discord).
• Providing reservoirs of stored energy for evolutionary processes: these processes are vital for maintaining order in a thermodynamic system tending toward entropy. Forests help maintain biosphere health and stability.
A review of selected literature
George Woodwell – ENERGY FLOW THROUGH THE BIOSPHERE
The Biosphere, Scientific American, 1970
Eugene Odum – DEVELOPMENT AND EVOLUTION OF THE ECOSYSTEM,
Chapter 9, Fundamentals of Ecology, Eugene Odum, 1971
• THE STRATEGY OF ECOSYSTEM DEVELOPMENT
• CONCEPT OF PRODUCTIVITY, Chapter 3, Fundamentals of Ecology, 1971
Howard Odum – SYSTEMS ECOLOGY: RESPIRATION – MAINTAINING INTERNAL ORDER AND STABILITY
Chapter 3, Fundamentals of Ecology, Eugene Odum, 1971
Paul Kreger – PHOTOSYNTHETIC ASSIMILATION, SYSTEM MAINTENANCE, and STABILITY
Energy Flow Through Watersheds, 1979
• Soils, Vegetation, and Climate Relationships
• Evaporative Cooling
• Forests Stabilize Micro and Macro Climates by Minimizing Soil Erosion with Layers of Leaf Litter and by Growing Interlocking Root Systems That Help Stabilize Seasonally Unstable Hills and Mountains
Erosion Processes in Steep Sloped Catchments, 1976 (Appendix A).
Robert Curry – ALL LIFE ON EARTH IS AN EXPRESSION OF ENERGY FLOW.
Energy flows from intense to less intense –
for example, from light to chemical, to motion, to heat – and at each transformation, a significant portion of energy
becomes heat waste. Watershed Form and Process, 1975
• Observations of thermodynamically efficient processes
Gregory Bateson – No organism, no mind, can know everything. Mind and Nature: A Necessary Unity, 1979
Wendell Berry – Excerpt from Kentucky River 1975 (Appendix B)
ENERGY FLOW THROUGH THE BIOSPHERE — Diversity, Stability, Efficiency
G. Woodwell, The Energy Cycle of the Biosphere, The Biosphere, Scientific American, 1970.
The energy that sustains all living systems (including human) is solar energy, fixed in photosynthesis and held briefly in the biosphere before it is reradiated into space as heat.
Photosynthesis is the process whereby sunlight, high intensity, shortwave energy, is absorbed by chlorophyll in green plants, combined with carbon dioxide from the air, hydrogen, and oxygen from water, and minerals from the soil, and converted to chemical energy in the form of carbohydrates.

This chemical energy becomes the roots, leaves, stem, branches, flowers, and fruits of the plant. And when the plant drops its leaves or bark, or dies and falls to the ground, it becomes the all-important organic component of the soil.
In combination with more photosynthesis in succeeding generations of plants this ever-increasing reservoir of organic energy slowly builds top soil, perhaps one inch every five hundred years. Occasionally, during volcanic eruptions, floods, or landslides, some of the organic material is buried in geologic substrates under the surface of the land or in the ocean floor and eventually through millions of years becomes coal and oil.
It is solar energy that moves the rabbit, the dog, the boy on the bicycle, my pencil, and my computer.
Solar energy has been fixed in one form or another on the earth throughout much of the earth’s 4.5 billion year history. The modern biosphere probably had its beginning about two billion years ago with the evolution of marine organisms that not only could fix solar energy in organic compounds but also did it by splitting the water molecule and releasing free oxygen.
The beginning was slow. Molecular oxygen released by marine plant cells accumulated for hundreds of millions of years, gradually building an atmosphere that screened out the most destructive of the sun’s rays and opened the land to habitation by living systems. The habitation of the land began perhaps 400 million years ago. New plant species evolved that were more efficient in absorbing light and converting it to chemical energy, thus accelerating the trend. This living process fitted the new species together in ways that not only conserved energy and the mineral nutrients utilized in life processes but also conserved the nutrients by recycling them, releasing more oxygen and making possible the fixation of more energy and the support of still more life. Gradually each landscape developed flora and fauna particularly adapted to that place. These new arrays of plants and animals used solar energy, nutrients, water and the resources of other living things to stabilize the environment, building the biosphere we know today.
In considering the fixation and flow of energy in ecosystems, it is important to consider the broad pattern of their development throughout the history of life on earth. If one were to ascribe a single objective to living processes, it would be the perpetuation of life. The entire strategy is focused on that single end. To achieve this end the resources of any location, including its input of energy, are divided among an ever-increasing number of different kinds of users, which we recognize as plant and animal species.
The arrangement of these species in today’s ecosystems is a comparatively recent event, and ecosystems continue to be developed by migration and continuing evolution. Changes occur slowly through a conjoint evolution that is not only biological but also chemical and physical. The entire life process appears to be open-ended, continuous, self-augmenting, self-healing and endlessly versatile. It builds on itself, not merely preserving life but increasing the capacity of a site to support life. In so doing it stabilizes the site and the biota. Mineral nutrients are no longer leached rapidly into watercourses; they are conserved and recirculated, offering opportunities for more life. This process is for all practical purposes stable. These are the living processes that have shaped the biosphere suitable for human habitation. These systems are self-regulating, self-healing and remarkably resilient.
The rich diversity and layering of plants and leaves in a relatively mature forest ecosystem have many species arranged in a structure that apportions the available solar energy as effectively and efficiently as possible. Forests, which cover about a tenth of the earth’s surface, fix almost half of the biosphere’s total energy (G. Woodwell, 1970).
DEVELOPMENT AND EVOLUTION OF THE ECOSYSTEM
Excerpts from E. Odum, Fundamentals of Ecology (3rd Ed.), Chapter 9, 1971
THE STRATEGY OF ECOSYSTEM DEVELOPMENT
Ecosystem development, of what is more often known as ecological succession, may be defined in terms of the following three parameters: (1) It is an orderly process of community development that involves changes in species structure and community processes with time; it is reasonably directional and, therefore, predictable. (2) It results from modification of the physical environment by the community; that is, succession is community-controlled even though the physical environment determines the patterns, the rate of change, and often set limits as to how far development can go. (3) It culminates in a stabilized ecosystem in which maximum biomass (or high information content) and symbiotic function between organisms are maintained per unit of available energy flow. The whole sequence of communities that replaces one another in a given area is called the sere; the relatively transitory communities are variously called serial stages or developmental stages or pioneer stages, while the terminal stabilized system is known as the climax. Species replacement in the sere occurs because populations tend to modify the physical environment, making conditions favorable for other populations until equilibrium between biotic and abiotic is achieved.
Succession involves a fundamental shift in energy flows as increasing energy is used for maintenance and system stability.
In a word, the “strategy” of succession as a short-term process is the same s the “strategy” of the long-term evolutionary development of the biosphere, namely, increased control of, or homeostasis with, the physical environment in the sense of achieving maximum protection from perturbations. The development of ecosystems has many parallels in the developmental biology of organisms, and also in the development of human society.
SYSTEMS ECOLOGY: RESPIRATION – MAINTAINING INTERNAL ORDER AND STABILITY
Excerpts from E. Odum, Fundamentals of Ecology (3rd Ed.) Chapters 3, 1971
Maintaining internal order is the number one priority in any complex system of the real world. The continual work of pumping out “disorder” is necessary if one wishes to maintain internal “order” and stability. (H. T. Odum, 1967, building on the concepts of E. Schrodinger, 1945, and A. J. Lotka, 1925). In any living organism, respiration is the process that pumps out disorder and maintains internal order. In the ecosystem, “order” in terms of a complex biomass structure is maintained by the total community respiration which continually pumps out disorder.
“Entropy” is the scientific term for disorder. Respiration pumps out entropy.
CONCEPT OF PRODUCTIVITY
Basic or primary productivity of an ecological system, community, or any part thereof, is defined as the rate at which radiant energy is stored by photosynthetic and chemosynthetic activity of producer organisms (chiefly green plants) in the form of organic substances which can he used as food materials. It is important to distinguish between Gross primary productivity and Net primary productivity. Gross primary productivity is the total rate of photosynthesis, including the organic matter used up in respiration during the measurement period. This is also known as “total photosynthesis” or “total assimilation.” Net primary productivity is the rate of storage of organic matter in plant tissues in excess of the respiratory utilization by the plants during the period of measurement. This is also called “apparent photosynthesis” or “net assimilation.” In all these definitions the term ”productivity” and the phrase ”rate of production” may be used interchangeably. Even when the term ”production” is used to designate an amount of accumulated organic matter, a time element is always assumed or understood, as, for example, a year when we speak of agricultural crop production. To avoid confusion, the time interval should always be stated.
The key word in the above definition is rate; the time element must be considered, that is, the amount of energy fixed in a given time. Biological productivity thus differs from “yield” in the chemical or industrial sense, in the latter case the reaction ends with the production of a given amount of material; in biological communities the process is continuous in time, so that it is necessary to designate a time unit; for example, the amount of food manufactured per day or per year.
In more general terms, the productivity of an ecosystem refers to its “richness.” While a rich or productive community may have a larger quantity of organisms than a less productive community, this is by no means always the case. Standing biomass or standing crop present at any given time should not be confused with productivity.
The overall relationships between GPP, Respiration (energy used for system maintenance and stability), and NPP can perhaps be explained by a graphic model.
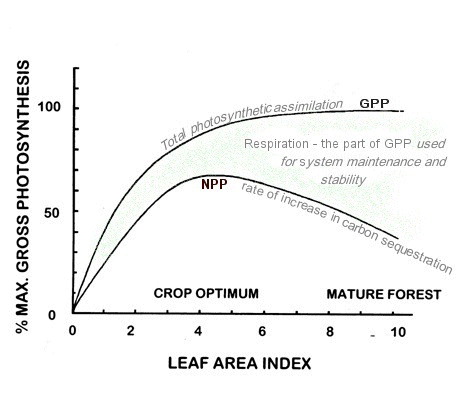
Leaf area index, as plotted along the x-axis of the graph, represents a measure of photosynthetic biomass. Maximum rate of net production is obtained when the leaf area index is about 4 (that is, the leaf surface exposed to light is four times the ground surface), but maximum gross production is reached when leaf area index reaches 8 to 10, which is the level found in mature forests.
Photosynthetic Assimilation, Carbon Sequestration, Respiration,
Entropy Pump Out, System Maintenance, and Stability
Excerpts from Paul Kreger, Energy Flow Through Watersheds,
California Resources Agency 1979
Some comments about the graphic model :
- Photosynthetic Assimilation (GPP) provides energy for growing roots, stems, branches, leaves, seeds & fruits (NPP). Photosynthetic assimilation also provides energy for system maintenance and stability (Respiration).
- Photosynthetic Assimilation builds on itself, not merely preserving life but increasing the capacity of a site to support life. In so doing it stabilizes the biological community and its geographical area.
- The GPP and NPP curves in the graphic model show rates of increase in productivity in relation to leaf area exposed to light per square centimeter of forest floor. The more leaves available to capture light, the better the productivity. A diversity of plants, some capturing direct light, and others indirect light, also improve productivity. The rich diversity and layering of plants and leaves in a relatively mature forest ecosystem have many species arranged in a structure that apportions the available solar energy as effectively and efficiently as possible.
- The NPP curve indicates that the rate of increase in production of standing biomass peaks at leaf area index of 4 and then slows. The accumulation of biomass continues, albeit more slowly, on to leaf area index 8 to 10.
- The gap between NPP and GPP is Respiration – energy used for system maintenance and stability. Respiration pumps out disorder – maintains internal order and efficiency.
- Intense light energy from the sun provides energy for photosynthesis, but only a small portion of the light hitting a leaf is used in photosynthesis. A significant portion of the intense energy is converted to heat at the surface of the leaf. The tree evaporates water through the leaves to prevent overheating, to maintain conditions optimum for photosynthesis, and for life.
- Photosynthesis in the leaf is a thermodynamic process. Solar energy is being transformed into carbohydrates. In all transformations of energy, a significant portion of the energy being transformed becomes low-grade heat waste.
- Water evaporating through leaves of trees cools the leaves, cools the forest, and recycles water across landscapes.
- Water recycling through forests is broadly correlated with the rate of GPP and associated respiration. Maximum recycling of water vapor occurs beyond leaf area index 8.
- Optimum biodiversity and community stability are attained above leaf area index of 9 or 10.
- Maximum storage of carbon in the biomass occurs beyond leaf area index 8.

- Natural forests have the genetic propensity to maximize photosynthetic assimilation, gross productivity, expansion of life, diversity and complexity, community stability – – durability, internal order, and thermodynamic efficiency.
- Forests, which cover about a tenth of the earth’s surface, fix almost half of the biosphere’s total energy (G. Woodwell, 1970).
SOILS, VEGETATION, AND CLIMATE RELATIONSHIPS
Soil is produced by the interaction of (1) vegetation and (2) climate on (3) the parent geologic material.
Distribution of vegetation is determined by (1) soil characteristics, such as organic content, water-holding abilities, nutrient content, and texture, and (2) climate.
Vegetation modifies the soil in which it grows (1) by building an organic or humus layer on top of the mineral soil, (2) by loosening the texture of the soil with the penetration of roots, and (3) providing microclimates suitable for mycorrhizae fungi. Networks of roots and fungi add organic content and air to the mineral soil thus increasing the nutrient content and water holding abilities of the soil. Vegetation adds stored energy to the soil.
Networks of strong roots help bind seasonally unstable slopes together. Layers of fallen leaves and decomposing organic materials shield the surface of the soil, minimizing surface erosion and small slippages.
Evaporative cooling through green leaves influences the temperature and moisture content of the soil and thus helps regulate its microclimate.
Vegetation influences climate (1) by recycling moisture into the air through the processes photosynthesis and evapotranspiration, which increases precipitation, and (2) by the moderation of temperature extremes, humidity, and wind.
Water circulates not only across the face of the earth and in the oceans deep but through life itself. The water that becomes a river begins as ocean vapor, drawn from the sea by the power of the sun and carried as clouds on the wind across the waves. These life-giving droplets of waters come to earth as mist and rain in forests. The water percolates into the humus rich floor of the forest, bringing the potential for birth and growth to all that it touches. Molecules of water are absorbed by mycorrhizae fungi and drawn to the roots of trees, shrubs and other forest plants — through the roots, up the stems, through branches to the leaves — photosynthesizing and respiring, evaporating and cooling the leaves of the trees, and re-humidifying the air. These molecules of water will again form mists and clouds and drift on the winds, condense again into droplets and fall to earth – cart-wheeling across valleys and mountains – recycling through vegetative photosynthesis / respiration three or four times before entering a stream and flowing / returning to the sea.
Change the type of vegetation which grows on and in the soil, or change the microclimate near the surface, and the organic, moisture and nutrient content and texture of the soil begins to change.
Changes in the distribution and type of vegetation or reduction of leaf surface area, over a large area will cause changes in the climate of that area and related areas.
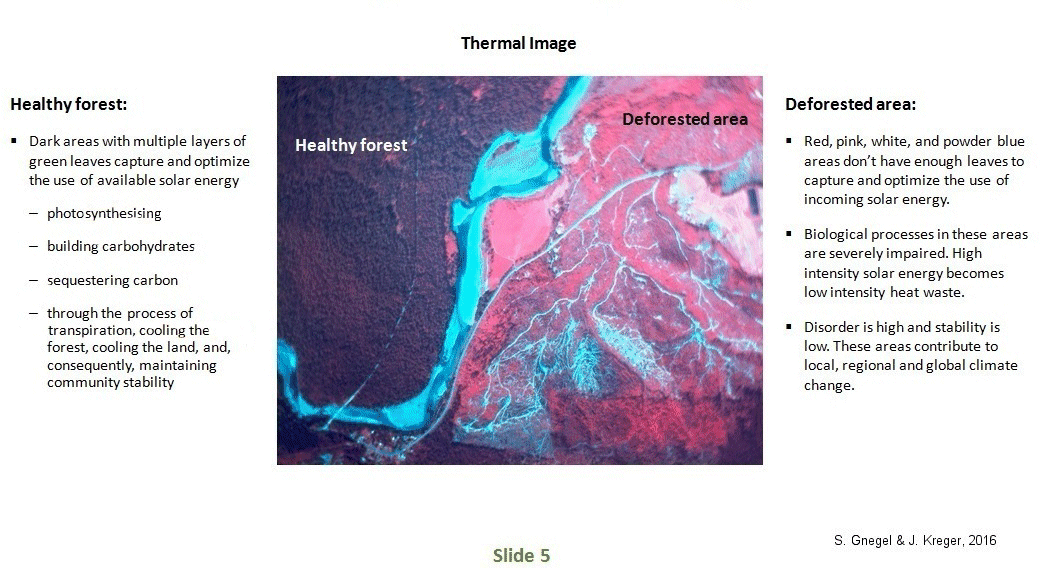
Climate and weather patterns over large land areas are influenced by the distribution and type of vegetation which lives on the land. The degree to which vegetation influences weather patterns is correlated to the total rate of photosynthesis.
The rich diversity and layering of plants and leaves in a relatively mature forest ecosystem has many species arranged in a structure that apportions the available solar energy as effectively and efficiently as possible. Climax forests in particular, because of their high leaf surface area to land ratio and high rates of respiration, are very important in weather cycles. Climax forests influence the wind, temperature, humidity, precipitation, soil, and water and energy flows in ways often discovered only after the forests are cut, and these functions are reduced or lost. Forests assist in the essential global recycling of water, oxygen, carbon, and nitrogen.
All life on earth is an expression of energy flow
Excerpts from R. R. Curry, Watershed Form and Process, 1975.
The energy that sustains all living systems (including human) is solar energy, fixed in photosynthesis and held briefly in the biosphere before it is reradiated into space as heat. Energy flows from intense to less intense. Every time energy is transformed from intense light to chemical, to motion, useable energy is decreased by the heat loss that occurs with each transfer of energy from one form to another. Each time we transform energy, we lose more than 90% of the applied energy to unusable heat waste. All systems, animate and inanimate, from grains of sand to microorganisms, and from watersheds to ecosystems, work within the natural patterns of thermodynamics.
Observations of “thermodynamically efficient processes” in natural systems reveal some fundamental patterns of behavior, action, reaction, and interaction:
- Sunlight photosynthesized by green plants powers life on earth.
- Plants absorb sunlight, store energy in the form of sequestered carbon, build communities, and strive to influence and stabilize their living space.
- These “natural systems” are self-regulating patterns of beings and forces, which take care of themselves and each other. [Self-regulating mechanisms are mechanisms which bring about a return to constancy if a system is caused to change from the stable state by a momentary outside influence.]
- Direct interactions (cell-to-cell, root-to-root, face-to-face exchanges within and across species) are those which enable the self-regulation of life to happen.
- Stability in the community is enhanced by the mutually supportive interactions of a diversity of beings.
- Matter and energy are moved around as little as possible–to minimize entropy (waste and disorder).
- Information is moved around as much as possible–to maximize synergy (efficiency and cooperation), to minimize entropy (waste and discord).
- Open and honest information flows – unobstructed feedback loops – are the processes that allow individuals and groups to self-regulate and self-correct.
- Open-flow feedback loops are necessary for maximizing potential benefits to the ecological community, and in achieving economic efficiency in the use of available solar energy and mineral resources.
- Organisms in the natural world run themselves in the most energy efficient way possible, with full “participatory flat power processes” and free flow of (honest) information. There are few (if any) remote-control, rule-from-afar, top-down hierarchical structures – they are simply not efficient.
No organism, no mind, can know everything or see the whole picture.
Gregory Bateson, Mind and Nature: A Necessary Unity, 1979
Free flow of information provides opportunities for many minds to focus on an issue and share their differing views, and thus develop larger mental descriptions and concepts. The greater the diversity of viewpoints and the more honest the information flow, the more complete the picture. Full-on competition of ideas and processes, continual feedback, expansion of mental processes, reduction of corruption, all enable continuous adjustment toward high efficiency / low entropy outcomes.
Summary
LIFE ON EARTH IS SUSTAINED BY THE FINITE AMOUNT OF SOLAR ENERGY ABSORBED, CONVERTED, STORED, AND RESPIRED BY GREEN PLANTS
Green plants are the “primary producers” of the biosphere, absorbing energy from the sun,
carbon dioxide from the air, water and minerals from the soil, and converting them into “energy rich” organic compounds that maintain the plants and all other living things. The process is called photosynthesis.
Chlorophyll in leaves
absorbs energy from the sun
carbon dioxide (CO2) from the air
plus water (H2O) from the soil
produces carbohydrates and releases oxygen.
C6 H12 O6 + 6O2
Photosynthetic Assimilation (Gross Primary Productivity / GPP)
provides energy for growing roots, stems, branches, leaves, seeds, and fruits (carbon sequestration / Net Primary Productivity / NPP).
Photosynthetic assimilation provides energy for system maintenance and stability (Respiration).
• Plant communities have the genetic propensity to maximize (within the constraints of their local environments) the rate of photosynthetic productivity (GPP).
• It is through maximizing GPP (including associated respiration) that plant communities exert stabilizing influences on earth’s life support systems, including biogeochemical cycles, and micro and macro climates.
• Without the stabilizing influences provided by strong plant communities, functioning at high rates of gross primary productivity, many of the biogeochemical cycles (carbon dioxide and water for example) become less stable, thus altering climate and threatening human food production and economic networks.
• The total rate of photosynthetic productivity (GPP) determines the total life possible on earth.
• On land, the total rate of photosynthetic productivity is closely related to the total leaf surface area exposed to light per unit of land. Leaf surface area is closely related to the height of the canopy, layering of leaves and diversity of species with varying requirements for light.
• Mature forest ecosystems typically have a rich diversity with many species arranged in a structure that apportions the available solar energy as effectively as possible.
• Photosynthetic assimilation builds on itself, stabilizing the biological community and the geographical area it lives in, expanding the opportunities for more life.
• Forests cover about a tenth of the earth’s surface and fixed almost half of the earth’s total photosynthetic energy (Woodwell, 1970).
• Since the early 70s accelerated logging, land clearing and burning have reduced the flow of photosynthetic energy to our life support system. More than two-thirds of the earth’s forests have been degraded.
• Water recycling through forests is broadly correlated with the rate of productivity. The more leaf surface area, the more GPP, the more respiration, the more water recycled (Odum, 1971).
We can optimize fiber production, or system stability, but not both at the same time. If we chose to manage
forests for maximum fiber production, we sacrifice biological and climate stability.



Appendix A
Forests Stabilize Micro and Macro Climates by Minimizing Soil Erosion with Layers of Leaf Litter
and by Growing Interlocking Root Systems That Help Stabilize Seasonally Unstable Hills and Mountains
Erosion Processes in Steep Sloped Catchments
P. Kreger, 1976
Introduction
Surface erosion occurs in the form of sheet and gully erosion on barren areas of mountain slopes. Sheet erosion is the removal of thin layers of surface material by storm runoff. It results from minute, numerous, localized pulses of running water flowing overland or from the impact of falling rain. Splash erosion is the dislodgment and movement of soil particles under the impact of falling raindrops. Gravity pulls the splashed particles down the slope. Surface erosion removes topsoil. Topsoil contains most of the organic material and nutrients necessary for growing forest trees.
Generally, natural undisturbed forest-covered slopes experience minimal loss of topsoil to surface erosion. Carpets of organic litter, decaying leaves, and small branches protect the soil from the erosive impact of falling rain. Organic litter tends to hold the rain where it falls. Organic litter provides habit for bugs and critters that make the soil more porous thus improving percolation of water into substrate layers. Organic litter helps maintain a relatively even distribution of water through the soil and across the slopes of the mountains, and thus slows the downhill movement of water. By reducing erosion, increasing percolation, and slowing the delivery of fast flowing water to streams, organic litter reduces the energy available for gully and stream channel erosion. The reduction of eroded earthen material and bed load sediments in a stream or river channel also reduce the potential for heavy flooding and downriver damage to life and property.
The geologic processes of uplift, folding, and fracturing, and downcutting by streams and rivers results in localized steep and unstable mountain slopes prone to large-scale mass erosion.
Steep slopes with weak structural or bedding planes, which lie nearly parallel to the angle of the slope, tend to fail more than slopes in which the structural planes are nearly perpendicular to the slope. Steep slopes with no internal structure are seriously unstable.
Slope failure processes in temperate forest regions are often associated with rainstorms, which temporarily load the soil matrix with detained water. During periods of prolonged precipitation, rain lubricates soil particles and decreases slope cohesiveness. Water increases the weight of the soil mantle, and gravity tends to pull the mass downslope. Expanding lattice clays at shear zones within the soil mantle increase the frequency of slope failures.
Roots of forest trees bind and hold steep and periodically unstable mountain slopes in place. If the roots die, the unstable slopes fail.
Landslides are potentially very destructive, causing significant losses in soil productivity, timber production, wildlife habitat, water quality, and fishery resources. Landslides also increase the probability of severe flooding, damage to property, and loss of life.
Clear cutting and road construction on unstable and marginally stable (metastable) slopes have been identified as major contributors to increased erosion and landsliding.
Landsliding
A landslide is the downward movement of a portion of a mountain slope. Landslides come in all sizes ranging from a few square meters in surface area to several square kilometres; from a meter or so deep to tens of meters deep. For simplicity, I will classify landslides as either (1) deep-seated, or (2) shallow. Deep-seated slides are more than four meters in depth. Shallow slides are usually less than four meters deep.
Deep-seated slides are frequently seen as “slope creep.” Slope creep is the slow (usually a centimetre or so per year) downhill movement of a large part of a mountainside or hill slope. Downward creep of mountain slopes occurs during times of prolonged and heavy precipitation when the earthen material of the slopes is saturated with water. During periods of moderate to dry weather, these slopes may appear to be relatively stable.
Steep sloping mountains in high rainfall areas have gullies and streams. At the bottom of the slopes are larger streams or rivers. During periods of heavy rainfall and high-stream flows the water (and the mud and gravel carried in the water) slowly cut down through the geologic material of the mountains. As the streams cut downward they undercut the toes of the slopes adjacent to the streams. In this way the slopes directly adjacent to, on either side and uphill of the streams become over-steepened. If the parent geologic material is unconsolidated or highly fractured these over-steepened slopes become shallow landslides. We call these steep, shallow landslides “debris slides.”
Debris slides occur on slopes that are at or near the angle of repose of the parent geologic material. They may be relatively dormant, supported and held in place by the roots of trees, or, wherever the streams are rapidly undercutting the slopes, the slides are active, fast moving, and with only light vegetative cover. Over-steepened and debris-slide slopes adjacent to and uphill of a stream may extend a few meters up the slope next to small ephemeral or intermittent stream, and from tens to a hundred meters or more along larger streams and rivers.
Over-steepened and debris slide slopes adjacent to streams are the toes of downward creeping mountain slopes. As the mountain slopes creep downhill, the toes of slopes are pushed into the streams. In other words, during seasons of high rainfall, the stream in flood flow is undercutting the base of the slope, the water in the mountainside increases the mass weight, lubricates the slope, and gravity pulls the slope downhill.
Landslides are often very complex. For example, near where I live, we have small landslides on larger landslides, shallow landslides on deep-seated landslides, and small active slides on larger dormant slides.
Although a slope may fail massively, the failure usually has small beginnings. A landslide, whatever its size, does not usually start in massive proportions.* It begins to fail at its weakest points. An unstable slope has many weak points both on its surface and underground. We might think of these weak points as triggers. Usually, these triggers are smaller unstable areas on or within the larger slope mass. These weak points are almost always associated with water in channels on the surface of the slope or water flowing through the slope mass. *The exception is when a sharp earthquake shakes a mountain slope laden with water. The slope momentarily loses cohesion and failure may occur in mass. This is called “liquefaction.”
During floods, streams exert tremendous forces on the stream banks, especially on the outside of curves in the channel. Almost all types of stream bank material will gradually wear away under this erosive force, but stream banks which consist of unconsolidated material as found on the face or at the base of active or dormant landslides erode very rapidly. Although stream bank erosion is a natural phenomenon, which occurs in all streams to some degree, it is a process that has been greatly accelerated by man’s activities.
Clearcutting and patch cutting are particularly damaging to soils and streams. Clearcutting and burning of logging slash frequently leave the soil mantle without organic cover; reduce the organic content and cause compaction of the soil. Soil without organic cover is compacted by the impact of falling rain. Rain falls at about 30 km per hour. In a moderate to heavy storm, raindrop impact amounts to more than 100 tons per acre per hour. The soil is also compacted where heavy equipment has been operated on the soil. Compaction of the soil reduces the amount of water absorbed and stored in the soil. Compaction makes for more rapid delivery of storm runoff to gullies and streams. More rapid delivery causes greater volumes and velocities of water in the gullies and streams. And each time the velocity of the water flow is doubled; its ability to move soil and rocks increases by sixty-four times. In other words, doubling the velocity of a stream allows the water to move a particle sixty-four times larger. Flow velocities in streams are determined by three factors, 1) steepness of slope, 2) combined volume of water and eroded material in flow, and 3) the roughness of the channel.
Stream channels and slopes adjacent to gullies and streams are exceptionally sensitive to increases in velocity of water flows. Increased flow means increased velocity means increased energy for stream channel and stream bank erosion. All of this results in further destabilization of stream channels and the undercutting of adjacent toes of stream-side debris slides.
Soil and rocks eroded from stream banks and adjacent slopes become sedimentation in stream channels. Sedimentation input into streams from disturbed slopes frequently occurs during periods of heavy precipitation. Man-induced erosion of earthen material enters stream courses from road cuts and fills, overland flow from compacted skid trails, disturbed ephemeral gullies and streams, and stream-side landslides resulting from excessive channel erosion which undercuts the toes of unstable stream-side slopes.
During low to medium intensity storms, earthen material (sand and silt) eroded from slopes disturbed by road construction or logging enters steep gradient tributary streams. The sand and silt are transported in the water as long as the velocity of the streams is sufficient to carry it. When the steep gradient streams join with the larger streams of lesser gradient, the sand and silt begin to settle to the bottom.
The addition of increased amounts of sediment to stream water increases the volume and velocity of flow. Sedimentation increases the total volume of material (water and sediment) that must be transported by the stream.
Soil and rocks, eroded from upstream, fill the deep pools of the stream. The bottom of the channel becomes flatter and less deep, and the stream channel widens. This is called stream channel aggradation. Aggradation is the addition of so much sediment to a channel that pools are filled in and the bed of the channel rises. Aggradation occurs either when sediment particles are too large to be carried by the water flow, or when there are too many particles to be carried. A typical result of aggradation is a decrease in the visible water flow in the stream in the summer and fall because a significant percentage of the water is flowing through or under the gravel of the aggraded streambed. And thus, aggradation, by decreasing the surface water flow in summer, decreases the available living space and rearing habitat of aquatic organisms.
Aggradation changes the channel dimensions both by decreasing the depth and increasing the width of the channel through bank erosion. Bank erosion undercuts the toes of streamside debris slides and thereby causes secondary land sliding and increased sedimentation. What we have then is the potential for a self-feeding loop — what we call exponential disequilibrium of the stream channel and adjacent slopes.

Appendix B
Mountains and valleys keep the shape
of the escape of water.
Running water carves the watershed
in violence so long and slow it is gentle.
It hollows the long main stem of the valley,
draws its fingers down the tributaries and coves,
and does it all in little displacements of stone,
little shifts and slips, nudgings,
wedgings of soil, dissolving into the stream.
And all the while the forest has been standing
dense and unmoved on the slopes,
and across the broad valley floor.
Trees and shrubs grow along the water’s edge,
Rainbow and Brown trout soar in pools
clear as the sky.
Millions of years it has been at this great work,
missing nothing. Meticulous and patient.
It works as a loving mind,
the fathering and mothering mind of God,
teaching its creation how to grow.
In flood it brings particles of humus
from the forest floor, and leaves it
on the bottomlands downstream,
richness to make myth behind us,
so great it is.
It is old light the currents carry.
Sunlight collected by the trees over hundreds
of thousands and millions of years
and put into the soil
to make the world a better place to live.
This old sunlight, old time itself,
the past, more vital and complete
than myth or memory, laid down
to bless the Earth.
from Wendell Berry’s 1975

Comments
How Do Forests Influence and Stabilize Climate? — No Comments
HTML tags allowed in your comment: <a href="" title=""> <abbr title=""> <acronym title=""> <b> <blockquote cite=""> <cite> <code> <del datetime=""> <em> <i> <q cite=""> <s> <strike> <strong>